1. INTRODUCCIÓN Y TEORÍA CELULAR
El conocimiento de la célula es reciente porque no es visible al ojo humano y no se pudo conocer hasta que no se obtuvieron las técnicas de observación. De hecho el conocimiento de la célula está unido al avance de las técnicas de observación. Esto comienza en el s.XV cuando un comerciante francés inventó un juguete para adultos basado en el principio de lentes convergentes en 1590, que ampliaban el campo visual. Este fue el primer microscopio.
En el siglo XVII su capacidad de observación estaba mediatizada por sus ideas pseudocientíficas religiosas. De todas formas, el primero que llegó en realidad al concepto de célula fue Hocke en 1665, que observando láminas de corcho con el microscopio vio unas cavidades poliédricas, que en realidad eran huecos celulares, y a esos huecos les llamó célula.
Todo estó provocó una serie de descubrimientos como el citoplasma, el núcleo...
el 1838 dos biólogos alemanes son los primeros en dar la teoría celular: La célula de Hoocke es la unidad estructural y funcional de los seres vivos capaz de valerse por sí misma, autónoma, capaces de realizar la nutrición relación y reproducción.
La célula es la unidad mínima de un ser vivo y se producen (en aquella época) por un impulso divino, llamado generación espontánea.
Virchow explicó el origen de la célula en 1858: Toda célula procede de otra célula. Por tanto la célula es la unidad anatómica y funcional de los seres vivos, autónoma y cuya procedencia viene de otra.
2. NIVELES DE ORGANIZACIÓN, ORIGEN Y ESTRUCTURA GENERAL DE LAS CÉLULAS
2.1 Niveles de organización
En el nivel molecular hay niveles de organización de los seres vivos. La materia orgánica tiene gran complejidad, como las proteínas, nucleótidos... Las macromoléculas no son un nivel biótico, sino que se asocian formando complejos supramoleculares, como los ribosomas...
Solo la célula tiene suficiente complejidad para ser autónoma y forma el primer nivel biótico.
Hay tres tipos de organización:
- Los virus: Son organizaciones bióticas, pero no celular. Son muy sencillos: Un complejo supramolecular en proteínas, ác. nucleicos... No tienen metabolismo, no tienen nutrición y no hacen la reproducción. Por lo tanto no son autónomos, son parásitos obligados. Lo único que saben hacer es la relación cuando detectan al tipo de célula a la que va a infectar. Es una forma de organización no celular y todo el resto de los seres vivos tienen un tipo de organización que es el celular.
- Los procariotas son células primitivas y sencillas, las bacterias.
- Las eucariotas son más complejas.

2.2. Origen y estructura general de la célula
El origen de la primera célula fue hace 3500x10e6 años. La Tierra se habría enfriado suficiente para que se formaran los océanos, una atmósfera más reductora (NH3, CH4, H2O...) que procedía de las erupciones volcánicas .
En estas condiciones, espontáneamente se habían originado los monómeros orgánicos, que caían al océano donde apareció la primera célula, pero antes debían aparecer los polímeros y los complejos supramoleculares.La primera célula aparece cuando se asociaron ácidos nucleicos capaces de expresarse en procariotas que realizan reacciones químicas, el metabolismo. Esto es la primera bacteria procariota.
 Esta primera bacteria procariota era fermentativa, no necesitan oxígeno, el problema surgió cuando terminaron las condiciones que permitían la formación de monómero, el alimento para esta célula empezó a escasear. Esto puso en marcha la selección natural favoreciendo a unas bacterias capaces de fabricar su alimento y permitió la perpetuación de la vida por la aparición de las plantas.
Esta primera bacteria procariota era fermentativa, no necesitan oxígeno, el problema surgió cuando terminaron las condiciones que permitían la formación de monómero, el alimento para esta célula empezó a escasear. Esto puso en marcha la selección natural favoreciendo a unas bacterias capaces de fabricar su alimento y permitió la perpetuación de la vida por la aparición de las plantas.
La célula eucariota apareció hace 1500x10e6 años. Fue originada por la fusión de los núcleos de células procariotas y también aparecen así los orgánulos membranosos. Más tarde esta célula "eucariota" tiene la capacidad de fagocitar (una célula se come a otra) y de esta manera aparecen las mitocondrias por fagocitosis de un procariota heterótrofo aerobio. Es la teoría endosimbiótica, que también explica la aparición de los cloroplastos y mitocondrias, que antes fueron procariotas autónomos que eligieron el camino de la simbiosis.
2.3 Estructura general de la célula
La célula procariota es muy sencilla, pequeña y primitiva. Todas las células procariotas tienen pared celular que no tiene celulosa, tiene una membrana plasmática y algunos poseen flagelos. El único orgánulo que tienen son los ribosomas para la síntesis de proteínas. Su información hereditaria forma ADN circular, con un solo cromosoma circular en posición central, que no está envuelto en la membrana nuclear, sino que está en el citoplasma donde hay algo más oscuro y denso, que es el nucleoide.
En la célula eucariota vemos más complejidad, mayor tamaño. La diferencia fundamental es la compartimentación que son los orgánulos membranosos que hacen que aumenten las funciones porque aumenta su tamaño. La ventaja de esto es que permiten realizar muchas funciones diferentes al mismo tiempo y es más eficaz, por tanto son células que necesitan menos materia y más energía.
Tienen:
- Ribosomas 80S, su ADN con las proteínas forma la cromatina, que forma los cromosomas.
- Dentro del núcleo se encuentra el nucleolo, que se encarga de la síntesis de ARNr y todo esto está envuelto en una membrana nuclear que es lo que sustituye el núcleo.
- El retículo endoplasmático que se encarga de la síntesis de lípidos y proteínas
- El aparato de Golgi que se encarga de la síntesis de glúcidos y el tratamiento de la secreción o almacenamiento de las sustancias del retículo endoplasmático. También se encarga de formar los lisosomas que tienen un papel fundamental en la digestión intracelular.
- Las mitocondrias cuya función es la respiración celular.
- El citoesqueleto, que se encarga del sostén de la célula. Una parte del citoesqueleto está relacionado con las células por su estructura celular.
- El centrosoma, que es un orgánulo estrellado que se encarga de los movimientos de los cromosomas durante la división celular.
- Las vacuolas que se encargan del almacenamiento de diversos tipos de cosas.
- Los cloroplastos, que realizan la fotosíntesis.

Diferencias entre célula vegetal y animal
Las células vegetales tienen:
- Pared celular celulósica
- Presentan cloroplastos, que contienen almidón o pigmentos fotosintéticos
- Tienen muchas vacuolas
- No tienen centrosoma
- Sí tienen huso acromático
- Las bacterias y las algas verdes azuladas son procariotas, los demás son eucariotas.
La diferenciación celular que hace que las células sean diferentes en estructuras, tejidos... es un proceso a partir de células embrionarias que son indeferenciadas y lo único que hacen es dividirse. En el desarrollo embrionario las células sufren la diferenciación celular que consiste en la represión irreversible del 90% del genoma de la célula.
3. MEMBRANA PLASMÁTICA
La aparición espontánea de la primera membrana fue determinante para la aparición de la primera célula. No se ve con el microscopio óptico y hasta hace poco no se pudo conocer su estructura. Para el estudio de la membrana se utilizan glóbulos rojos porque son semicélulas. La membrana de los glóbulos rojos está formada por un 40% de lípidos y un 60% de proteínas, que puede varias según sus funciones.
3.1 Lípidos
Los lípidos que forman la membrana son híbridos que tienen dos polos, uno polar y otro apolar. Esto se llama lípidos anfipáticos. Cuando están en un medio acuoso toman una forma de bicapa. Estos lípidos son fosfolípidos, pero también hay glucolípidos, glicerolípidos y colesterol, típicos de células animales.

Los lípidos determinan la estructura y propiedades de la membrana.
1) Una de las propiedades de la membrana es que se autoensamblan, se fusiona una membrana de una célula con otra, como con las pompas de jabón y se alargan las membranas por contacto sin perder la continuidad. Esta propiedad le permite a la célula la exocitosis (autosellado). La endocitosis es cuando la membrana se acorta sin perder continuidad debido a la afinidad química (anfipáticos) de sus lípidos. Esto permite que nunca haya un contacto directo, interior ni exterior de la célula. Esta propiedad es aprovechada en cosméticos o fármacos dentro de las células envueltas en vesículas lipídicas, son los liposomas.

2) Otra propiedad es el punto de fusión de los lípidos; La membrana son fosfolípidos que tienen ácidos grasos y como son cortos y saturados, presentan alto punto de fusión, por tanto es sólida y es inútil. Lo que ocurre que además de fosfolípidos, hay esteroides que tienen anillos muy insaturados, lo que hace que disminuye el punto de fusión y hace que las membranas sean fluidas También ocurre porque las fuerzas que unen las fuerzas de las membranas son débiles, ya que son interacciones hidrofóbicas, fuerzas de Van der Walls y a demás de fluidas son flexibles. Esto permite el movimiento de los lípidos por difusión, es lo que se llama 3) difusión lateral. La difusión de arriba a abajo nunca puede ocurrir porque se perdería la bicapa. Esto no ocurre gracias al colesterol, que estabiliza la bicapa.

4) Otra propiedad es su semipermeabilidad; Es permeabilidad selectiva, no es selectiva con las sustancias apolares, pero sí con las sustancias polares, que son más grandes. La membrana es apolar.
3.2 Proteínas
Las proteínas determinan su funcionamiento. El paso de las proteínas en la membrana depende de su afinidad con los lípidos. Son proteínas intrínsecas (hidrófobas) y se colocan atravesando la bicapa de fosfolípidos y quedando incrustada. Hay otras hidrófilas que no pueden atravesar la bicapa y se quedan unidas a las cabezas polares. Son proteínas periféricas. Las proteínas también tienen difusión igual que los lípidos pero a menos velocidad.

3.4 Modelo del mosaico fluido
La estructura de la membrana celular se dio en 1972 por Siger y Nicholson. Los lípidos y las proteínas de la membrana forman un mosaico, un puzzle fluído y flexible donde pueden difundir. Además, la membrana es asimétrica.
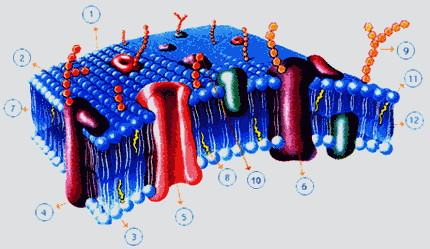
3.5 Funciones de la membrana plasmática
En la membrana ocurren una serie de reacciones químicas que son las que causan los intercambios de materia y de información entre la célula y el medio. Otras son las que producen los movimientos celulares.
Por ejemplo, los transportadores, que son proteínas, o los receptores hormonales, realizado por otro tipo de proteína. Hay dos sistemas de transmisión de información: El sistema nervioso y el sistema hormonal, mediante hormonas, que son como mensajeros químicos y que van por la sangre hasta las células dianas donde desencadenan unas reacciones (efectos que produce la hormona). Hay diferentes tipos de hormonas: Lipídicas que atraviesan la membrana y polares que no pasan la membrana. Estas últimas tienen receptores hormonales que ponen en movimiento a una enzima que transforma AMP y lo convierte en AMPc , que desencadena las reacciones de la hormona dentro de la célula. Provoca efectos como el cambio de la temperatura celular, aumenta la velocidad del metabolismo, activa la síntesis de ciertas proteínas, activa enzimas reguladoras, cambian el estado metabólico...
La enzima fosfodiesterasa, que convierte el AMPc en AMP, es decir, que destruye el transmisor inmediantamente, para que su efecto no sea continuo, inútil. La cafeína inhibe la fosfodiesterasa.
Otro ejemplo es la actividad ATPásica, consiste en ATP + H20 <--> ADP + Pi + Energía.
Hidroliza el ATP desprendiendo la energía para el transporte de sustancias o movimientos siempre asociados a proteínas contráctiles.
- TRANSPORTE DE PEQUEÑAS MOLÉCULAS: La membrana al ser semipermeable no puede ser atravesada por todas las sustancias, ya que tiene permeabilidad selectiva de las sustancias que la atraviesan. Hay dos formas: Transporte pasivo que es espontáneos y sin coste de energía y transporte activo que consume energía y la membrana, según qué transportador sea, elige cómo ser atravesada.
- Transporte pasivo: Un soluto se disuelve en agua por difusión hasta que se igualan sus concentraciones empujados por la presión osmótica. Esto es un gradiente químico, un movimiento por difusión. También puede haber una difusión por gradiente eléctrico. En los seres vivos se suelen dar gradientes electroquímicos a favor de gradiente de concentración. Las sustancias apolares atraviesan la membrana por difusión, afinidad química. Por ejemplo los ácidos grasos o las hormonas esteroideas, las sexuales, el oxígeno... porque son apolares. Las moléculas muy pequeñas, como le agua, atraviesan la membrana por los canales acuosos y los iones por los canales iónicos. Estos canales son proteínas intrínsecas, proteínas muy apolares que atraviesan la membrana. Esto es lo que llamamos difusión simple. Las moléculas polares grandes, como los monosacáridos o amoniácidos, necesitan el concurso de un transportador, que son proteínas de la membrana que se unen específicamente a la molécula. Esto es difusión facilitada. El transportador se une a la sustancia y cambia de estructura girando en la membrana, siempre a favor de gradiente.
- Transporte activo: Igual que la difusión facilitada pero el cambio de estructura del transportador consume energía y es el transporte que es en contra de gradiente o a mayor velocidad que la difusión facilitada. Esto ocurre para moléculas pequeñas.


- TRANSPORTE DE GRANDES MOLÉCULAS: El transporte de grandes moléculas, como los virus o las bacterias, tienen otros mecanismos específicos por endocitosis o exocitosis. (FIGURA 12.7)
4. PARED CELULAR
4.1 Estructura de la pared celular
La pared celular de los eucariotas es extracelular. Se origina a partir del aparato de Golgi por secreción desde la división celular, que es la citocinesis por tabicación en las células vegetales.
En la primera fase después de la división lo primero que aparece es la lámina media, que está formada de polisacáridos y glucoproteínas. Sobre esta lámina media se forman hasta tres capas de haces de celulosa (polisacáridos) paralelas entre sí dentro de cada capa y cruzados respecto de las capaz anteriores y posteriores, que es lo que le da la estructura y consistencia casi cristalina. También un cemento constituído de polisacáridos, hemicelulosa, proteínas y sales.
Esta es la pared celular primaria en la que predomina el cemento sobre la celulosa, pared que pertenece a las células vegetales embrionarias. Cuando ocurre la diferenciación celular es cuando se forma la segunda pared típica de las células maduras diferenciadas en tejidos, formada de 3 a 20 capas de celulosa sobre el cemento.
Ciertas células de ciertos tejidos presentan unas modificaciones en la pared celular, por ejemplo la sabia que forma los vasos del xilema acumula en su pared segundaria lignina, que es rígina y sostiene a la planta.
4.2 Funciones de la pared celular
La pared celular es rígida, entonces su función básica es que sostiene a la célula vegetal y no necesitan esqueleto. La otra función es la de proteger a las células vegetales del choque osmótico.
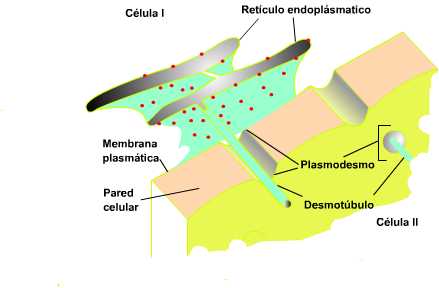 En la pared celular vegetal presentan unas perforaciones por donde contactan y se comunican las células a través de sus membranas plasmáticas. De este modo se intercambian las sustancias (plasmodemos). En otros casos pasan a través de canales membranosos compartiendo su retículo endoplasmático.
En la pared celular vegetal presentan unas perforaciones por donde contactan y se comunican las células a través de sus membranas plasmáticas. De este modo se intercambian las sustancias (plasmodemos). En otros casos pasan a través de canales membranosos compartiendo su retículo endoplasmático.
5. HIALOPLASMA SOLUBLE O HIALINO.
5.1 Composición
Es un líquido viscoso, una dispersión coloidal en estado de sol, citosol. Se encuentra entre la membrana plasmática y nuclear y es blanco y gelatinoso, estructurado por una red de proteínas que se llaman citoesqueleto, que es el medio celular.
Si al citosol le quitamos las proteínas del citoesqueleto aparecen inclusiones, que son gotas de lípidos, y lo que nos queda es el hialoplasma soluble que está formado por el 80Ç% de H20, monómeros, sales, metabolitos (sustancias intermedias del metabolismo).
También nos encontramos los ARNs (síntesis de proteínas) y enzimas que catalizan reacciones metabólicas del hialoplasma. El metabolismo del hialoplasma es el metabolismo intermediario. Por eso se dice que es una encrucijada metabólica donde también ocurre el catabolismo anaerobio de los azúcares, que son las fermentaciones, la forma más antigua de conseguir energía (sin O2) y se ha conservado por todas las células. Este proceso es llamado glucólisis.
En el hialoplasma también se da la conversión mecano química de la energía, donde se convierte el ATP en movimiento.
6. ORGÁNULOS NO MEMBRANOSOS
6.1 Citoesqueleto y estructuras afines
El citosol es gelatinoso, amorfo y blando pero no está suelto, sino que está perfectamente estructurado por una red de proteínas que sostienen a los orgánulos y da forma a la célula.
Forma parte de otras proteínas que originan otras estructuras celulares, cilios, flagelos, movimientos del cromosoma... y todas están relacionadas con movimientos celulares y son muy semejantes a los microfilamentos de las células musculares en su estructura, composición y funcionamiento, además de que todas se inhiben con la misma sustancia: citocolasina B.
- MODELO DE LOS FILAMENTOS DESLIZANTES: Es el modelo que explica el funcionamiento de las células musculares. Presentan una estriación y tiene sarcómeros , que es la unidad estructural y funcional de las células musculares. Un sarcómero está formado por dos tipos de proteínas: miosina que son filamentos gruesos. En los filamentos gruesos hay otros filamentos más delgados y son de actina. Los filamentos finos tienen unas cabezas y cuando el músculo se contrae consume energía que se invierte en unir las cabezas de la miosina a los filamentos delgados (actina) provocando el deslizamiento de los filamentos gruesos fijos sobre los delgados. Los sarcómeros se acortan, no los filamentos. Tienen actividad ATPásica intrínseca. Hay tres tipos de filamentos:
- Microfilamentos: Son los más delgados y está formados por una proteína globular, la actina, que polimeriza y forma como una especie de cuerda de dos hebras en hélice. Son los que producen el movimiento de la membrana por acortamiento y alargamiento a través de la fagocitosis, endocitosis y exocitosis.
- Filamentos intermedios: Son variados y dependen de los distintos tipos celulares. Por ejemplo, las neurofibrillas o las fibras de queratina de las células epidérmicas.
- Microtúbulos: Son filamentos más gruesos formados por tubulina. Están organizados de distintas maneras que constituyen las base de sustentación del sitoesqueleto, el huso acromático, los centriolos, los corpúsculos basales, los cilios y flagelos. os microtúbulos tienen una característica y es que tienen polaridad, se polimeriza y se despolimeriza. Son estructuras bastante complejas que se montan y se desmontan con bastante facilidad según sus necesidades celulares. Otros, como los centriolos, cilios y flagelos son más estables que el huso acromático.
- Centro organizador de microtúbulos: Todo estas estructuras se forman en centro organizados de los microtúbulos en la célula, como el que está en el centrosoma, que es un orgánulo pequeño en forma de estrella que se encuentra en la periferia del núcleo. Es exclusivo de células animales. La forma estrellada se debe a unos microtúbulos que salen llamados aster. En el centro hay dos cilindros llamados centriolos que se encuentran perpendiculares uno al otro. Uno está formado por 9 triplomicrotúbulos. La sustancia que rodea a los centriolos se llama sustancia pericentriolar, ese es el centro organizados de microtúbulos polares del huso acromático.
- Cilios y flagelos
- Estructura: Los presentan las células animales y células vegetales. Son aparentemente muy diferentes, ya que los cilios son muchos pelillos cortos y producen un movimiento de remado. Además son muy numerosos. Los flagelos se presentan como uno o dos y se mueven por propulsión. Tienen la misma estructura y funciones. Son orgánulos recubiertos por una membrana que es una continuación de la membrana plasmática. Están formados de "cañas", son como cilindros y es un microtúbulo formado por 9 duplomicrotúbulos. De esos 9 duplomicrotúbulos, sólo uno continúa hasta el corpúsculo basal, que también es un cilindro formado por 9 triplomicrotúbulos. Uno de ellos se continúan con el del flagelo o cilio. Estos y el corpúsculo basal son interconvertibles entre ellos. En el corpúsculo basal existe en la parte próxima (cara que da al flagelo o cilio) un cilindro central de proteínas que es al que se únen los 9 microtúbulos. Es la llamada rueda de carro. El corpúsculo basal es muy parecido al centriolo, tanto que son interconvertibles.
- Funcionamiento: Los microtúbulos de cada uno de los duplomicrotúbulos se encuentran próximos entre sí y entre ellos hay una proteína, la dineína, parecida a la miosina, que tiene actividad ATPásica intrínseca, capaz de hidrolizar ATP sin una enxima, se debe a ella misma (es una propiedad de la proteína). Esto le permite convertir la energía química del ATP en energía mecánica provocando el desplazamiento de los microtúbulos (conversión mecano química de la energía) igual que en las células musculares. Esto produce el deslizamiento del microtúbulo flotante sobre el microtúbulo fijo (continuo). En el cilio, cuando se deslizan se produce la flexión produciendo así que los cilios puedan remas. En los flagelos se produce una onda de flexión en la parte inferior, que produce un movimiento ondulatorio de propulsión.


Son los únicos orgánulos macizos de las células. Están formados por dos subunidades distintas fisiológicamente, de tamaño, composición y de coeficiente de sedimentación, que se calcula con un tubo de ensayo donde se observa a la velocidad que cae.
Todos los ribosomas están formados por el 60% de H2O y el peso seco de procariotas 70s es de un 60% de ARNr y un 40% de proteínas. Estas son 55 proteínas distintas.
La subunidad grande es de 50s y la pequeña de 30s aunque juntas tienen un coeficiente de sedimentación menos que la suma de las dos subunidades por independiente.
El de las eucariotas es 80s. Es más complejo pero semejante y el peso seco es un 50% de ARNr y un 50% de proteínas. Las subunidades son 60s y 50s. Tiene 88 proteínas distintas. El ARNr es monocatenario y aunque tiene una alta proporción (60% de sus nucleótidos) son complementarios dentro de la misma cadena. En este 60% se forma una doble hélice dentro de la misma cadena (estructura secundaria). El 40% restante forma bucles, sumados forman la estructura terciaria. Esto hace que sea compacta y le da la consistencia maciza.
Los ribosomas se forman en los nucleolos. El centrómero es lo que une las cromátidas hermanas en los cromosomas y a veces presentan un segundo centrómero en el que hay genes que cuando se transcriben sirven para fabricar los ARNr. El nucleolo es la cromátida con centrómeros unidos más el ARN polimerasa más los nucleótidos de ARN que da lugar a la transcripción de los ARNr. Pero también se ensamblan los ARN del nucleolo y de otros cromosomas más las 55 proteínas (metabolismo postransquipcional).
De esta forma se producen las subunidades por separado y salen al hialoplasma donde autoensamblan en torno a un ARNm y empieza la síntesis de proteínas.
Los ribosomas aparecen en todas las células porque es el único orgánulo esencial, excepto en los espermatozoides maduros. Los ribosomas los encontramos libres y aislados en los procariotas, igual que en los eucariotas solo que aquí no están aislados, sino que forman polisomas y también presentan ribosomas asociados a la membrana (RER.Mn) donde sintetizan proteínas libres asociadas a membranas.
En cuanto a su funcionamiento, son los realizadores mecano químicos de las proteínas. Su funcionamiento comienza cuando autoensamblan por estereoespecifidad. Son complementarios y aparecen propiedades nuevas cuando ambas subunidades están unidas que por separado no tienen. Estas propiedades consisten en que interaccionan reversiblemente con otras moléculas con los factores de iniciación, elongación y terminación (síntesis de proteínas). La llegada y salida de estas moléculas produce un cambio en la estructura de todas sus moléculas y en la estructura del ribosoma completo que desemboca en la síntesis de proteínas.

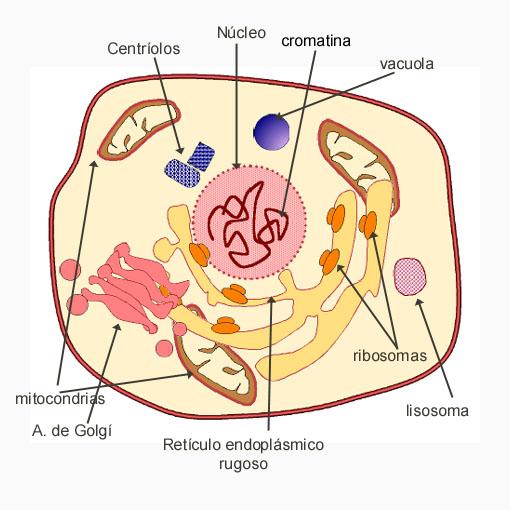
7. ORGÁNULOS MEMBRANOSOS
Las células eucariotas poseen un complejo de orgánulos membranosos que no tienen las procariotas. Esto sería el resultado de la primera célula eucariota como fusión de célula procariota y se originaría el núcleo y otros orgánulos membranosos como explica la teoría endosimbiótica. Es lo que produce una compartimentación que permitió a los eucariotas aumentar la velocidad de su metabolismo y además se consigue realizar más funciones al estar compartimentadas.
7.1 Retículo endoplásmico
Es un laberinto membranoso que ocupa el 10% del volumen celular. Es una sola membrana muy plegada. Se divide en dos: El RE liso porque no tiene ribosomas y el RE rugoso porque su cara externa presenta ribosomas. Su función (RER) es la síntesis de proteínas que la célula quiere almacenar dentro que son exportadas o almacenadas. El RE liso se encarga de la síntesis de los lípidos y de combinar lípidos con glúcidos, formando así el glucocáliz.
El RE en las células musculares es abundante y se llama sarcoplasmático, donde almacenan iones Ca++ y los expulsa. Supone una señal para la contracción muscular.
El RE tiene relación directa con la membrana nuclear, es la continuación de la parte especializada del RER y además tiene una relación indirecta con la membrana plasmática a través de su relación con el aparato de Golgi.
7.2 Aparato de Golgi
Está formado por un conjunto de vesículas membranosas, dicoidales, aplanadas y apiladas. Es como una pila de platos llamados dictiosomas y el conjuntos de dictiosoma es el aparato de Golgi.
 El aparato de Golgi es abundante en las células secretoras, son glandulares. Está relacionado con el retículo endoplasmático, cuyas sustancias transforma y empaqueta para su almacenamiento posterior o secreción. Tras la fabricación de las sustancias en el RE lo abandonan en vesículas de transcripción hasta llegar a la cara de formación más cercana (sáculo). Va pasando por cada sáculo hasta llegar a la cara de maduración donde las sustancias se abandonan en vesículas de secreción saliendo del aparato de Golgi para ser almacenada o para atravesar la membrana plasmática.
El aparato de Golgi es abundante en las células secretoras, son glandulares. Está relacionado con el retículo endoplasmático, cuyas sustancias transforma y empaqueta para su almacenamiento posterior o secreción. Tras la fabricación de las sustancias en el RE lo abandonan en vesículas de transcripción hasta llegar a la cara de formación más cercana (sáculo). Va pasando por cada sáculo hasta llegar a la cara de maduración donde las sustancias se abandonan en vesículas de secreción saliendo del aparato de Golgi para ser almacenada o para atravesar la membrana plasmática.
7.3 Lisosomas
 Son vesículas membranosas que miden entre 0,3 - 1,5 mm. que contienen hasta 40 enzimas digestivas (hidrolasas ácidas). La células se digiere y se fabrica el RER, AG Y REL. Esto sirve para la digestión intracelular.
Son vesículas membranosas que miden entre 0,3 - 1,5 mm. que contienen hasta 40 enzimas digestivas (hidrolasas ácidas). La células se digiere y se fabrica el RER, AG Y REL. Esto sirve para la digestión intracelular.
Lo presentan todas las células. Es autofagia porque se regeneran, típico de los organismos unicelulares. Pero en pluricelulares no ocurre esto. Se llama heterofagia. Sus células reciben los monómeros a través de la sangre, que es digestión extracelular.
7.4 Vacuolas

Son vesículas membranosas. Las tienen todas las células y las vegetales tienen más. Las vacuolas tienen como función almacenar sustancias de reserva, proteínas, toxinas, pigmentos, h2o que penetra por ósmosis en la vacuola manteniendo la turgencia de las células vegetales para aumentar la superficie aumentando el intercambio entre células.
7.5 Mitocondrias
Son orgánulos aislados. Sin embargo con un microscopio fluorescente se puede observar que se encuentran unidas unas con otras.
Es un orgánulo cilíndrico, tiene doble membrana.
La célula eucariota anaerobia ancestral estaba por heterofagia a punto de digerir a un procariota primitivo aerobio cuando escuchó una vocecita que dijo: ¡No me comas, no me comas, si me das monómeros yo te fabrico tela tela de ATP que te mola y necesitas tanto! Entonces el eucariota vio que era bueno y establecieron una relación de colegueo a la que llamamos "relación interna de simbiosis" y fueron felices (y comieron...¿procariotas?) hasta el día de hoy.
Una membrana mitocondrial externa es lisa y más permeable, sobretodo a los iones y a las moléculas de bajo peso molecular.
La membrana interna tiene repliegues, que son crestas mitocondriales y que tienen como función aumentar la superficie impermeable (como sería una membrana plasmática)
 Otra característica es que es una membrana plasmática sin colesterol, solo tiene en las células procariotas. Tiene un 80% de proteínas, son transportadores y presentan cadenas transportadoras de electrones.
Otra característica es que es una membrana plasmática sin colesterol, solo tiene en las células procariotas. Tiene un 80% de proteínas, son transportadores y presentan cadenas transportadoras de electrones.
Entre las membranas hay un espacio intermembranoso, como un "hialoplasma". Dentro de la mitocondria se encuentra la matriz mitocondrial, que tiene en su interior un "hialoplasma" que tiene H2O, iones, monómeros, ADN circular (plásmidos) que contiene información para fabricar transportadores. También tiene genes para formar las enzimas para su propia duplicación. Las mitocondrias se reproducen ellas solas y todas son de origen materno. La genética evolutiva estudia el ADN mitocondrial que es pequeño, clónico y prácticamente idéntico al que hace 10e6 años porque son libres de combinación.
La matriz mitocondrial presenta muchas enzimas para duplicar, transcribir, sintetizar sus proteínas y los ribosomas son 70s.
Además también tiene enzimas para el catabolismo aerobio que ocurre aquí, que son transportados por la célula hospedadora. Esto es la simbiosis.
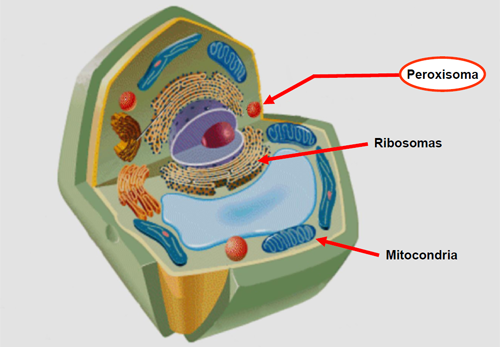 Son parecidos a los lisosomas. En su interior ocurren reacciones de oxidación y detoxificación. Lo hace la enzima catasa y eliminan también el etanol. Abundan en el hígado y riñón que se encargan de su eliminación.
Son parecidos a los lisosomas. En su interior ocurren reacciones de oxidación y detoxificación. Lo hace la enzima catasa y eliminan también el etanol. Abundan en el hígado y riñón que se encargan de su eliminación.7.7 Cloroplastos.



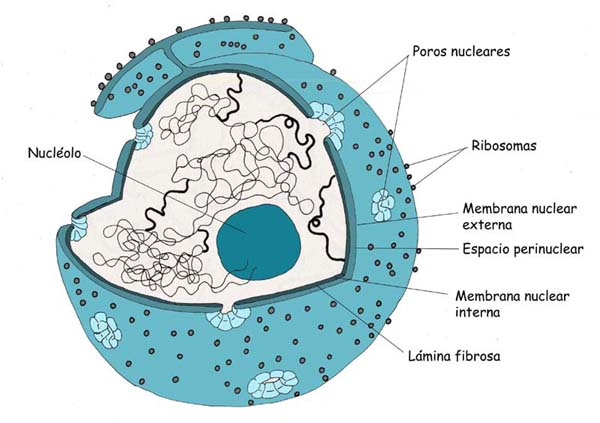
7.8 Núcleo
No hay comentarios:
Publicar un comentario